Introduction
The relationship between vegetation ecosystems (the compositional, structural, and landscape characteristics) and the dispersal and evolution of hominins has long been a major focus in palaeoecology, archaeology, palaeoanthropology, and related disciplines1,2. The early Pleistocene was a critical period for global climatic and environmental changes, characterised by cooling and aridification trends, as well as for the migration and dispersal of Homo erectus3,4,5,6. Hominin dispersal first occurred from Africa to Eurasia7,8,9. While existing studies have predominantly emphasised the climatic drivers of hominin evolution10,11,12,13, emerging evidence underscores the crucial role of vegetation ecosystems in shaping dispersal corridors and adaptation strategies14,15,16,17,18. The open savanna vegetation of Africa is widely regarded as having facilitated the evolution and dispersal of H. erectus19,20. Nevertheless, the climatic patterns and vegetation types of Eurasia diverge markedly from those of Africa, rendering the African model difficult to apply to Eurasia21. Specifically, Eurasia may exhibit distinct patterns of vegetation landscapes and hominin dispersal evolution.
Over the past decades, a series of innovative studies have been conducted on the relationship between hominin evolution and vegetation landscapes in Eurasia2,17,21. This has resulted in two different hypotheses: (i) the open habitat hypothesis, in which hominins were better adapted to occupying open steppe or forest-steppe ecological environments21,22, where the open vegetation landscape provided favourable conditions for the migration, hunting, and survival of early H. erectus17; (ii) the relatively closed habitat hypothesis, in which the environmental characteristics during the period when H. erectus was active were forest vegetation landscapes23,24. This forest vegetation not only provided rich food sources for hominins but also offered safe tree-dwelling habitats23,25. However, there have been no detailed studies on to which the vegetation landscape was open to be suitable for the migration, hunting and survival of early hominins.
The main reasons for this debate and the lack of in-depth research are: (i) Issues, including discontinuous sedimentation or the poor preservation of sedimentary strata, have resulted in limited information about the changes in past vegetation ecosystems near archaeological sites16,26. (ii) Most studies are limited to a single artefact layer or a relatively short time span, and have a low temporal resolution, making it difficult to comprehensively reconstruct vegetation ecosystem processes before and after the activities of H. erectus27,28. (iii) Most research is based on qualitative analysis, and there is a lack of systematic quantitative studies of the impact of vegetation ecology and landscape characteristics on the evolution of H. erectus29. Therefore, to better understand the influence of early Pleistocene vegetation composition and landscape openness on the migration and evolution of early hominins, it is necessary to conduct quantitative research on paleovegetation and landscape openness, based on stratigraphic sequences with continuous sedimentation, and employing multiple artefact layers, sensitive proxy indicators, a high temporal resolution, and covering a long interval.
Pollen analysis plays an irreplaceable role in revealing the paleovegetation and past landscapes openness30,31. Early studies relied on non-arboreal pollen (NAP) percentages to infer vegetation openness32, but this approach fails to account for pollen productivity variations and dispersal biases, resulting in vegetation–pollen mismatches33. Subsequent semi-quantitative alternatives, such as the arboreal/non-arboreal pollen ratio (AP/NAP)34 and the difference between the maximum score of forest biomes and that of open biomes within the biome classification, have yielded improved insights but lacked precise quantification35. Application of the Regional Estimates of Vegetation Abundance from Large Sites (REVEALS) model has resulted in significant research progress by incorporating pollen productivity corrections and nonlinear vegetation–pollen relationships to quantify openness, its exclusion of the bare land proportion has remained a constraint33. To address these limitations, our study employs a transfer function methodology to establish pollen assemblage–vegetation openness relationships. This approach enhances the reconstruction reliability by utilising entire pollen spectra rather than individual taxa to minimise single-type interpretation biases36.
The Nihewan Basin in northern China has the largest and most concentrated group of early Pleistocene Palaeolithic sites in East Asia37. More than 280 Palaeolithic sites have been found in this region, of which 23 are older than 1.0 Ma38,39 (Fig. 1a, b, Supplementary Table 1). Among them, the Majuangou (MJG) archaeological site is the earliest hominin site, and it is also the earliest stone-flake tool site in Northeast Asia in terms of reliable stratigraphic relationships and abundant discovered materials40. Ten artefact layers have been identified at the MJG site, which provide a framework for the cultural sequence of hominins from 1.76 to 1.32 Ma, in the early Pleistocene39. Notably, the activity surface of a mammoth butchery site was revealed within the MJG–Ⅲ artefact layer (~ 1.66 Ma), and more than 60 mammoth footprints were identified in MJG–Ⅱ (~ 1.64 Ma) (Supplementary Fig. 1)39. This is the earliest and the richest artefact assemblage among the early Pleistocene sites in the Nihewan Basin40. In addition to archaeological research, environmental change research has also been conducted at the MJG site37. Qualitative studies have been conducted on several artefact layers, including of mineral composition, ostracods, and animal fossils, which have revealed the burial environment and fossil animal assemblages of the artefact layers37,41,42. However, quantitative studies are lacking and several key questions have not been addressed, including: What was the relationship between hominin activities and vegetation landscape openness? What degree of vegetation openness was conducive to hominin activities?
Fig. 1: Location and field photos of the MJG archaeological site in the Nihewan Basin, China.
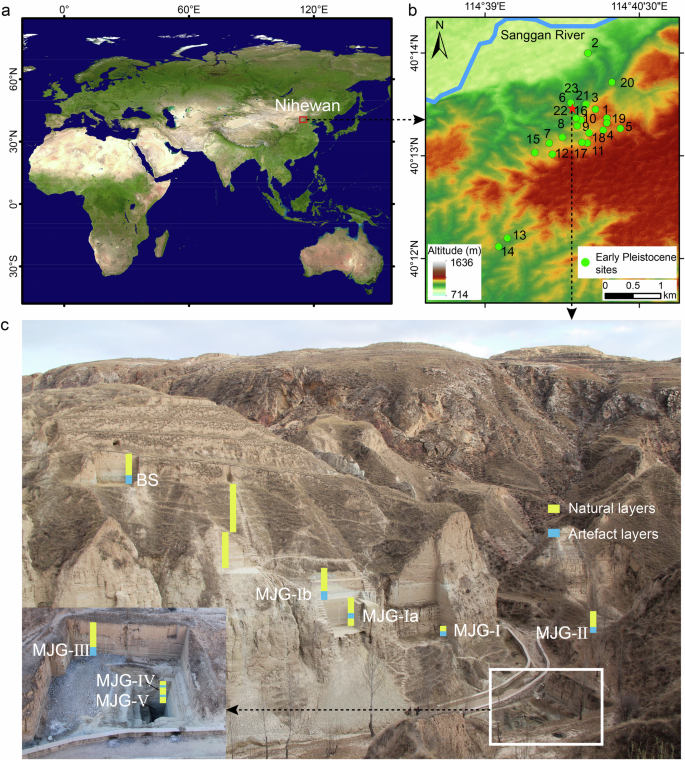
a Location of the Nihewan Basin (map source: https://search.earthdata.nasa.gov/) (Supplementary Note 1). b Distribution of sites over 1 Ma in age in the Nihewan Basin. For detailed information about sites 1–23, see Supplementary Table 1 (map source: https://www.gscloud.cn/). c Site profile and artefact layers of the MJG site in the Nihewan Basin, from Northwest to Southeast (image by Fagang Wang).
In this study, based on the existing paleomagnetic chronology and 26Al/10Be burial dating37,43, ages of the eight artefact layers in the section of the MJG site (40°13′31.066″N, 114°39′50.493″E, 832.6 m a.s.l) were calibrated using the sedimentation rate (Fig. 1c). We conducted pollen analysis on 422 samples collected at 10-cm intervals from eight artefact layers and natural sedimentary layers. The REVEALS model33 and transfer function methods36 were then utilised to quantitatively reconstruct the changes in vegetation cover and landscape openness during and around the period of hominin activity in the Nihewan Basin. This study aims to reveal which types of vegetation landscape and openness degree in the early Pleistocene of the Nihewan Basin were more conducive to attracting hominin activities. Overall, we hope to provide detailed and reliable evidence for the study of the impact of paleoenvironmental changes on the migration and evolution of hominins during the early Pleistocene in China and even the world.
Results and discussion
Stratigraphy and chronology of the MJG archaeological site
The MJG archaeological site is located in the northeastern Nihewan Basin, northern China40,44 (Fig. 1a, b). Due to the submersion of the bottom two artefact layers by groundwater during sampling, we selected eight artefact layers and their surrounding natural layers for analysis40. Sedimentological analysis, incorporating grain size metrics and field stratigraphic evidence, indicates that the studied profile consists predominantly of silt (average of 70%), which is consistent with the characteristic depositional of the Nihewan Formation38,45 (Fig. 2, Supplementary Note 2).
Fig. 2: Lithology and chronology of the sedimentary profile at the MJG archaeological site.
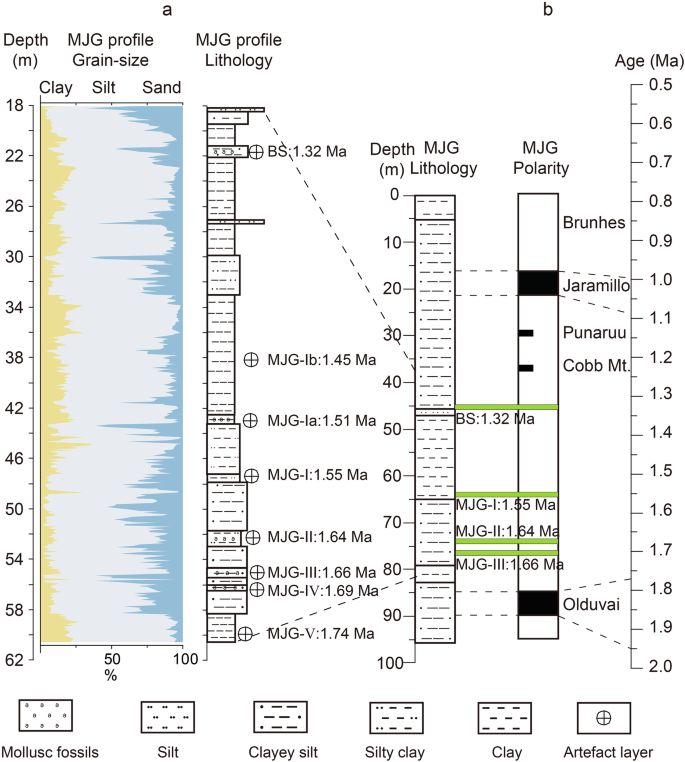
a Lithology and grain size composition of the MJG profile (Supplementary Note 2, Supplementary Data 1). b Lithology and chronology of the MJG profile37.
The lithology of the MJG profile from bottom to top is as follows (Fig. 2): 60.5–58.1 m, grey and greyish-brown clayey silt with horizontal bedding; 58.1–42.5 m, grey and greyish-yellow clayey silt; 42.5–18 m, greyish-brown and grey silty clay with horizontal bedding (Supplementary Note 1). Samples of the MJG profile were collected continuously from the bottom upwards at 10-cm intervals, with some samples taken at 12-cm intervals. The total sampled interval was 60.5–18 m, with a thickness of ~42.5 m. A total of 422 samples were collected.
The chronology of the MJG profile was well constrained by magnetostratigraphy37 and 26Al/10Be burial dating43. In 2004, Zhu et al.37 established the magnetostratigraphic framework for the profile (Fig. 2b), confirming that the BS, MJG–Ⅰ, MJG–Ⅱ, and MJG–Ⅲ artefact layers reside in the Matuyama reverse chron, bracketed by the Olduvai and Jaramillo normal subchrons37. Based on average sedimentation rates, their ages were derived as: BS (1.32 Ma), MJG–Ⅰ (1.55 Ma), MJG–Ⅱ (1.64 Ma), and MJG–Ⅲ (1.66 Ma)37. In 2008, extrapolated ages for MJG–Ⅳ (1.69 Ma) and MJG–Ⅴ (1.74 Ma) were derived using these chronologies and sedimentation rates (Fig. 2a)44. In 2024, Tu et al.43 applied 26Al/10Be burial dating to BS, MJG–Ⅰ, MJG–Ⅱ, and MJG–Ⅲ layers, with results concordant with Zhu et al.'s magnetostratigraphic ages, providing robust cross-validated chronological constraints.
Two additional artefact layers (MJG–Ia and MJG–Ib) discovered in 2013 occur stratigraphically above MJG–Ⅰ40. The MJG–Ia layer is located 4.5 m above the MJG–Ⅰ, while MJG–Ib layer is ~4.2 m above MJG–Ia and ~16.8 m below BS (Fig. 2, Supplementary Note 1 and Note 2)40. Their ages were determined through grain size variations (Fig. 2a): The average sedimentation rate ( 11 cm/ka) between MJG–Ⅰ and BS was calculated from stratigraphic separation and age difference. Sediments between MJG–Ⅰ and MJG–Ia show coarser grain size, while MJG–Ia to MJG–Ib intervals are finer, with a grain size difference factor of ~1.35. Based on these parameters and sedimentation rates, MJG–Ia and MJG–Ib were dated to 1.51 Ma and 1.45 Ma, respectively.
Quantification of modern vegetation landscape openness
Vegetation landscape openness, a key component of landscape ecology, is utilised in modern vegetation research to gauge the cover of non-arboreal vegetation (e.g., grasses and shrubs). This index ranges from 0 (representing 100% arboreal canopy closure, as in a dense forest) to 100% (indicating the complete absence of woody vegetation, characteristic of open grassland). Empirical studies demonstrate an inverse relationship between arboreal cover and openness: lower arboreal plant coverage rates correspond to higher vegetation landscape openness values46.
In this study, we first assessed the consistency between the reconstructed and modern observed vegetation landscape openness. To achieve this, we conducted field validation across 57 systematically selected sites spanning a gradient from closed forest to open grassland within the study region (see Methods, Supplementary Note 4). The vegetation landscape openness quantification utilised MOD 44B Version 6 Vegetation Continuous Fields product (250 m spatial resolution), accessed via NASA’s Earthdata platform (https://search.earthdata.nasa.gov)47. This dataset quantifies three land-cover components: % tree cover, % non-tree cover, and % non-vegetated cover. For this analysis, vegetation landscape openness was calculated as:
Vegetation landscape openness = 100%−% Tree cover
Application of this model showed that 85.9% of the reconstructed vegetation landscape openness samples were within the error range (Fig. 3a). The change trend between the reconstructed and observed values was also relatively consistent, with the correlation coefficient (_R_2) of 0.87 (p < 0.05) (Fig. 3b). The reconstructed vegetation landscape openness was consistent with the observed values, indicating that the model was dependable. This showed that this technique could then be applied to the stratigraphic data to reconstruct temporal changes in the openness of the vegetation landscape at the MJG site during the early Pleistocene.
Fig. 3: Comparison of observed and reconstructed vegetation landscape openness in modern surface samples.
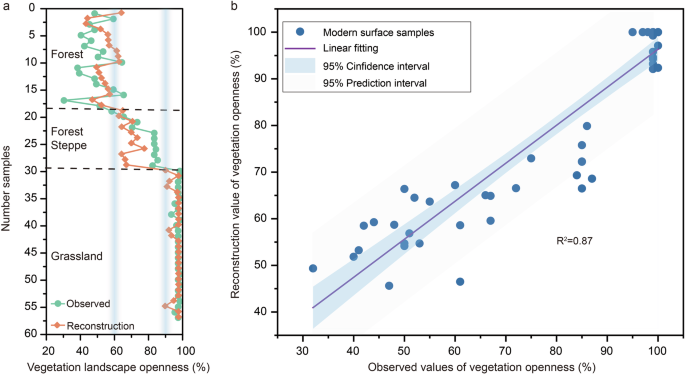
a Sample numbers 1–18 correspond to forest vegetation, 19–29 to forest-steppe, and 30–57 to grassland (see Methods, Supplementary Data 2). The figure shows observed and reconstructed vegetation landscape openness values across three vegetation types: forest, forest-steppe, and grassland. The two light blue columns represent the critical errors for forest/forest-steppe and forest-steppe/grassland. b Correlation coefficient between observed and reconstructed openness values for forest, forest-steppe, and grassland vegetation landscape openness.
Vegetation cover and landscape openness of the MJG site
Prior to quantitatively reconstructing vegetation landscape openness at the MJG site, we compared the performance of different distances, models, and methods (Supplementary Table 2). The comparative analysis revealed that a 1000-km radius modern pollen dataset achieved the highest predictive accuracy (_R_2 = 0.82, Fig. S3), which was incorporated into the quantitative reconstruction of vegetation openness at the MJG site. Subsequently, we tested the degree of matching and its statistical significance between the fossil pollen assemblage of the MJG profile and modern pollen assemblage (see Methods, Supplementary Fig. 4). The MJG profile fossil pollen samples exhibit a high degree of correspondence with the regional modern pollen assemblage (Supplementary Fig. 4a), and the results of vegetation landscape openness reconstructions have statistical significance (p < 0.05) (Supplementary Fig. 4b). The vegetation landscape openness reconstruction outcomes are consistent with independent vegetation cover estimates (excluding bare ground) and the biome type derived from pollen assemblages (Fig. 4, Supplementary Fig. 5b, Supplementary Note 5; Supplementary Tables 3 and 4). These comparisons confirm the reliability of our quantitative reconstruction of vegetation landscape openness and vegetation cover. This result is supported by independent global climate records, including the LR04 benthic δ¹⁸O stack48 and the magnetic susceptibility record of the Lingtai loess section in China49 (Supplementary Fig. 5e, f). This consistency with independent multi-proxy records strengthens our confidence in the accuracy of our vegetation cover and landscape openness reconstruction.
Fig. 4: Changes in vegetation cover and landscape openness during the early Pleistocene at the MJG site.
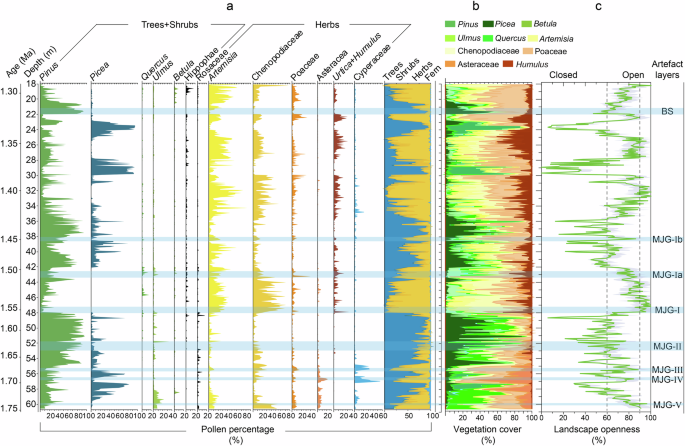
a Percentages of the principal pollen types. b Vegetation cover reconstructed based on the REVEALS model. c The openness of the vegetation landscape was quantitatively reconstructed based on the transfer function method (Supplementary Note 4, Supplementary data 3).
The vegetation landscape openness of the eight artefact layers of the MJG site during the early Pleistocene is mainly between 60% and 90% (Fig. 4). The average vegetation landscape openness for each artefact layers were as follows: MJG–Ⅴ (average of 88.3% and the same below), MJG–Ⅳ (66.4%), MJG–Ⅲ (78.7%), MJG–Ⅱ (58.5%), MJG–Ⅰ (91.9%), MJG–Ia (78%), MJG–Ib (65.9%), and BS (60.8%).
Based on vegetation composition, the eight artefact layers could be divided into two main types (Fig. 4). The first group consists of six artefact layers (MJG–Ⅴ, MJG–Ⅳ, MJG–Ⅲ, MJG–Ⅰ, MJG–Ia, and MJG–Ib), dominated by Artemisia, Chenopodiaceae, and Poaceae, and the herb pollen content is >50%, while the tree pollen content is relatively low, <30%. This suggests a forest-steppe vegetation landscape in the study area. The openness of these six layers (>65%, up to 90%) further supports their classification as forest-steppe (Fig. 3). The second group comprises two artefact layers (MJG–Ⅱ, and BS), dominated by Pinus, with the tree pollen content up to 80%, while the herb pollen content was ~20% on average. The vegetation landscape openness at this was ~60%, which indicates an open temperate forest vegetation landscape.
However, when vegetation openness was <50% or >90%, there was minimal or no evidence of hominin activities. Where Picea pollen percentages exceeded 30% (e.g., in the natural layers located at the upper part of MJG–Ⅴ, MJG–Ⅳ, MJG–Ⅱ, and MJG–Ib) and vegetation openness was <50% (even <20% in some layers), it indicates that the vegetation landscape was a cold temperate coniferous forest. When herbaceous plants consistently accounted for a high proportion (>90%), accompanied by vegetation openness exceeding 90% (e.g., in the natural layers located at depths of 47.2–43.8 m (1.55–1.52 Ma), 33.8–31.3 m (1.41–1.39 Ma), and 27.7–24.8 m (~1.36–1.34 Ma)), indicating a steppe environment. In these layers, neither the steppe environments characterised by persistently excessive vegetation openness nor the forest environments with relatively high canopy density constituted the most favourable habitats for hominin.
In summary, through the comparison between the natural layers and the artefact layers, openness of the vegetation landscape reconstructed for the eight artefact layers is mainly between 60% and 90%, which indicates that a relatively open vegetation landscape was more conducive to the migration, hunting, and survival of hominins in the Nihewan Basin.
Relatively open vegetation landscape in the early Pleistocene promoted the evolution of early hominins
This study provides compelling evidence for hominin adaptation to a relatively open vegetation landscape. The openness of the vegetation landscape reconstructed from the eight artefact layers was mainly between 60% and 90%. In contrast, when the openness of the vegetation landscape was <50% or >90%, there was no or very little evidence of hominin activities. This comparison indicates that hominins selectively occupied the Nihewan Basin during their dispersal process, where relatively open vegetation landscapes facilitated their dispersal.
To investigate whether this linkage between a relatively open vegetation landscape and hominin activity extended beyond the Nihewan Basin, we compiled all the published data from other early Pleistocene archaeological sites in Eurasia. A total of 24 archaeological sites from the early Pleistocene period (1.8–1.0 Ma) across Eurasia were collected in our study (Supplementary Data 4). Although the majority of these prior studies provided qualitative methods, we performed a statistical analysis of the available data, using the vegetation descriptions provided by the original authors (e.g., forest, forest-steppe, and grassland)50 (Fig. 5a). Forest vegetation landscapes are categorised as closed vegetation landscapes, while forest-steppe and grassland are categorised as open vegetation landscapes. Completely open grassland vegetation landscapes, defined as those lacking any tree cover, were excluded from the final statistical comparison.
Fig. 5: Location and vegetation landscape of global archaeological sites for the interval of 1.8–1.0 Ma.

a Global distribution of 1.8**–**1.0 Ma archaeological sites and the proportion of open and closed vegetation landscapes in Eurasia (archaeological sites are listed in Supplementary data 4). b Pattern diagram of open and closed vegetation landscapes. Credit: elements such as trees, herbs, hominins, and animals are from Huaban.com. Drawing on both these elements and the insights from this study, I reconceived and drew this diagram.
The results show that >83.3% of Nihewan archaeological sites, 77.8% of China's archaeological sites, and 73.9% of Eurasian archaeological sites had a dominantly relatively open vegetation landscape during the period of early Pleistocene hominin activity51,52,53 (Supplementary Fig. 5a). A comprehensive study also supports the inference that a relatively open vegetation landscape in the early Pleistocene was conducive to hominin evolution and migration26. This common pan-Eurasian pattern suggests that a relatively open vegetation landscape was critical for the dispersal and adaptive success of H. erectus (Fig. 5b).
Interestingly, this preference for relatively open vegetation landscapes was not unique to Eurasia1,54,55. Evidence from Africa also indicates a similar environmental preference among hominins. In northern Kenya’s Turkana Basin, carbon isotope analysis (δ13CVPDB) of pedogenic carbonates from the early Pleistocene suggests that hominins inhabited savanna ecosystems with ~40% woody cover and >60% open vegetation54. In the Awash and Omo-Turkana Basins of eastern Africa, using stable carbon isotopes analyses from the late Pliocene epoch, combined with 13C/12C ratios from 1300 palaeosols in adjacent areas, further indicate that woody cover was at most sites below ~40%, reinforcing the prevalence of open landscapes in hominin habitats1. In east and south African, a comparative analysis of Plio-Pleistocene mammalian fossil assemblages, alongside 31 extant mammalian communities from eight different habitat types, reveals that Homo was the first hominid to occupy fairly open, arid grasslands55. Across Africa, environmental data from 22 early Pleistocene archaeological sites consistently depict a predominantly open tropical savanna landscape as the primary background for early hominin evolution (Supplementary Data 4).
Conversely, relatively closed forest vegetated areas might have impeded the migration56 and visibility of H. erectus, while long-term open grassland vegetation landscape areas could have been unfavourable for hominin survival due to arid climate, scarce food resources, and lack of habitat57. This ecological constraint may explain the relative scarcity of early Pleistocene (1.8–1.0 Ma) archaeological sites in both low-latitude dense forest regions and mid-to-high latitude arid grassland zones (Fig. 5). It was precisely these vegetation characteristics, either excessively closed or overly open, that might have restricted hominin dispersal.
In summary, our findings, which are corroborated by evidence from multiple regions worldwide, indicate that relatively open vegetative landscapes were key to facilitating the migration and dispersal of early Pleistocene hominins. This pattern appears to be a prevalent and global phenomenon, deeply intertwined with the ecological preferences and adaptive strategies of H. erectus.
Mechanisms of hominin evolution in a relatively open vegetation landscape
During the early Pleistocene, relatively open vegetation landscapes likely played a significant role in facilitating the early hominin evolution17,52. These landscapes provided an increased abundance of animal and plant resources, enhanced visibility for resource acquisition, and reduced the risk of predation, all of which favoured ecological adaptation and cultural innovations17. Our study demonstrated that such relatively open vegetation landscapes exhibited a high Simpson’s diversity index (generally >0.6) and species richness, with a diverse vegetation composition, which provided diverse and abundant plant resources for early hominins (Supplementary Fig. 5c, d, Supplementary Note 3).
Moreover, sedimentological evidence indicates that the hominin occupation layers are associated with coarse-grained, lakeside facies deposits58 (Fig. 2), which would have ensured sustained access to water resources for early hominins44,53. These lakeshore environments not only promoted the growth of herbaceous plants, creating relatively open vegetation landscapes that attracted herbivores intensively exploited by hominins for hunting14,59, but also provided an abundance of local lithic raw materials1,38,60,61. The availability of these resources enhanced hominins’ ability to exploit both animal and plant resources, thereby promoting the development of stone tool technology, as evidenced by the increased core exploitation efficiency within the MJG–Ib artefact layer40.
The animal and plant fossils discovered in the artefact layers at the MJG site support our conclusions in this study38. Specifically, fossils of steppe animals including rhinoceros, deer, and horse have been discovered in the MJG–V, MJG–IV MJG–Ia, and MJG–Ib artefact layers40,62 (Supplementary Note 1). Abundant mollusk shells (including Gyraulus chihliensis and Planorbis youngi), and the leaves and seeds of aquatic plants were found in MJG–Ⅲ, suggesting a lakeside or swamp environment suitable for large herbivores and carnivores38. A substantial quantity of fossils of Mammuthus trogontherii, exhibiting signs of smashing and scraping, were discovered in MJG–Ⅲ (Supplementary Fig. 1). These fossils indicate scenes of hominins dismembering animals, scraping bones for meat consumption, and utilising bones, and they confirm that large animals were a crucial food source for these hominins38 (Supplementary Fig. 1). The analysis of mammal fossils from the MJG–Ⅲ artefact layer by Qiu et al.62 revealed that steppe animals accounted for ~70%, while forest animals accounted for ~15%. More than 60 intact elephant footprints were uncovered in the MJG–Ⅱ artefact layer (Supplementary Fig. 1). The analysis of biological fossils recovered from the BS layer revealed elephantids, cervids, and rhinocerotids that were adapted to a forest environment, while Equus was adapted to a grassland environment38,63. This evidence indicates that, in addition to forest, a significant area or areas of grassland existed within the regional vegetation during the period of hominin activity64. Qiu et al.62 analysed animal fossils in the BS layer and found that steppe animals accounted for ~60% of the total. Fossil evidence from the MJG site demonstrate that during the period of hominin activity, the relatively open vegetation landscape offered a diverse range of resources. The co-occurrence of fossils from both forest-adapted and grassland-adapted animals, coupled with evidence of hominin utilisation of large animals, suggests that this mixed vegetation setting was conducive to hominins survival.
Notably, similar scenarios highlighting the importance of relatively open vegetation landscapes for hominins are also evident at multiple early Pleistocene H. erectus archaeological sites around the world65,66,67. In northwestern Kenya in Africa, data from 481 fossil tracks, including 97 hominin footprints attributed to H. erectus, reveal that the open vegetation landscape of the lakeshore provided hominins with water resources, abundant food resources (including aquatic and terrestrial animals and plants), offering efficient hunting/scavenging opportunities, and enhanced visibility that facilitated migration65. Similarly, the open vegetation landscapes in Java, Indonesia, and at Kocabaş in the Denizli Basin (Southwestern Turkey) attracted large numbers of big herbivores, providing H. erectus with potential animal resources for survival66,67.
In summary, our results suggest that a relatively open vegetation landscape facilitated the evolution and migration of early H. erectus, while dense forest vegetation hindered hominin dispersal and technological development. Although climate played an important role in the migration and evolution of early hominins68,69,70, the influence of vegetation landscape openness cannot be ignored2,17. Overall, our findings offer a novel perspective on human–environment interactions during the period of hominin evolution, emphasising the role of the vegetation landscape as a key selective pressure alongside climatic change.
Methods
Pollen analysis
Pollen analysis was performed at 10-cm intervals on 422 samples, with a sample temporal resolution of approximately 1000 years. Sample preparation followed the conventional HCl–NaOH–HF treatment71. For each sample, 300 g of sediment was weighed and one tablet of Lycopodium spores (27,560 grains) was added to calculate the pollen concentrations. After chemical treatment, pollen and spores were extracted using heavy liquid flotation. These procedures were conducted at the School of Geographical Sciences of Hebei Normal University. Pollen identifications were made at ×400 under a Zeiss Imager A2 optical microscope with the aid of standard pollen reference publications for China72 and reference material preserved in the Key Laboratory of Environmental Evolution and Ecological Construction of Hebei Normal University. A minimum of 400 terrestrial pollen grains was counted for each sample. The common pollen types are illustrated in Supplementary Fig. 6. Pollen diagrams were drawn with Tilia 1.7.1673.
Quantitative reconstruction of vegetation landscape openness
Sources of modern pollen and vegetation landscape openness data
The modern pollen dataset we used is based on China’s modern pollen database, comprising a total of 4164 samples74 (Supplementary Fig. 2). The data on vegetation landscape openness corresponding to the modern sampling points are MODIS satellite data provided by the National Aeronautics and Space Administration (NASA), with a spatial resolution of 250 m (https://search.earthdata.nasa.gov/).
Screening the modern pollen data
Screening of modern pollen data is crucial for ensuring the accuracy of quantitative reconstructions. The selection process we applied to the modern pollen dataset employed the following criteria: 1) Air trap and dust samples were excluded due to differences in the preservation of modern pollen compared to fossil pollen. 2) Samples with latitude, longitude, and altitude that did not match the real situation were excluded (a total of 18 samples were excluded in this study). 3) Fan et al.75, based on modern pollen data and the Human Influence Index (HII, 0–64) (https://sedac.ciesin.columbia.edu/)76, successfully distinguished the threshold values of native/secondary vegetation and secondary/anthropogenic vegetation by using the error inflection point–discriminant technique (EIPDT) (HII = 22 and 38, respectively). Vegetation begins to be affected by human activities when HII > 22; therefore, modern pollen samples with HII > 22 were deleted in this study.
In the study, the pollen assemblage was dominated by temperate taxa, with only trace amounts of subtropical and frigid zone types detected (Fig. 4). This composition suggests that the past vegetation likely developed under a temperate climatic regime. Consequently, to accurately reconstruct past vegetation landscape openness, it is crucial to select modern pollen data from a comparable spatial range. Given the dominance of temperate pollen and this regional climatic context, the space-for-time substitution approach was applied. The selection of an appropriate spatial range for the modern training set is therefore critical for model performance77. An excessively small range may not capture the full variation within the reconstruction sequence, while an excessively large range can weaken the pollen-landscape relationship and introduce errors77. Based on principles of quantitative reconstruction, we established modern pollen training sets within multiple distance intervals (800–1500 km from the study site, at 200-km intervals) for analysis77. Among the tested intervals, the training set within a 1000 km radius yielded the optimal model performance (Supplementary Table 2). This optimal dataset comprised 1201 modern pollen samples (Supplementary Fig. 2). It covers a wide gradient of vegetation landscape openness (27%–100%) and includes all the principal pollen assemblage taxa found in the fossil record (Fig. 4).
Quantitative reconstruction of vegetation landscape openness
We selected the three most widely used methods for the quantitative reconstruction of vegetation openness: the Weighted Averaging Partial Least Sequence (WA-PLS), the Modern Analogue Technique (MAT)36 and Random Forest (RF)78. These three methods are currently the most widely used79,80. We compared the results obtained using these three methods to select the most suitable method in this study. The predictive performances of all the calibration models were assessed via self-cross-validation, with reference to the performance statistics of each calibration model, including the root mean square error of prediction and _R_2 between observed and predicted values36 (Supplementary Fig. 3, Supplementary Table 2). We also used non-metric multi-dimensional scaling to test the degree of matching between modern and fossil pollen assemblages81, and the random TF method was used to evaluate the statistical significance of the pollen and landscape openness reconstructions81.
First, to verify the reliability of the reconstruction results of the chosen model, we selected 57 modern samples from the transition between forest and grassland in the vicinity of the study area (the Taihang mountains, Guancen mountains, Nihewan, and Zhangbei regions in North China), which covered the major pollen assemblages within the strata (Supplementary Note 4). Then, the vegetation landscape openness was reconstructed for two purposes: to verify the correlation between the reconstructed and observed values, and to determine the critical values of the vegetation openness of forest, forest-steppe, and grassland (Fig. 3). All these analyses were conducted using R (4.0.3) with the “rioja” (version 1.0–7) and “palaeoSig” (version 2.0–7) packages.
Quantitative reconstruction of vegetation cover
The Landscape Reconstruction Algorithm (LRA) is based on the relative pollen productivity of the dominant plant taxa in a given study area, and the relative vegetation abundance is reconstructed using the REVEALS model33. The REVEALS model is intended for reconstructing regional vegetation using pollen records from large lakes (e.g., larger than 100 ha) or a combination of multiple small lakes33. It can correct the differences in pollen productivity and dispersal characteristics among taxa, while also accounting for the type and size of sedimentary basins33. This model has been widely applied in vegetation reconstruction from the late Pleistocene in Europe and Asia82,83,84. The results demonstrate that, compared to unadjusted pollen percentages, the plant abundances estimated by the REVEALS model more accurately reflect true regional vegetation coverage83. Previous studies have shown that the Nihewan paleo-lake began to form during the late Pliocene85. The paleo-lake covered an area of ~10,000 km² between 2.18 and 1.87 Ma, being the largest at the time85. From 1.87 to 0.73 Ma, it retreated by 3–4 km, reducing its area to an estimated ~8000 km285. Consequently, since this conforms to the underlying assumption of large lakes, the REVEALS model is appropriate for reconstructing past vegetation cover in this region.
The REVEALS model was run using the LRA. REVEALS.v6.2.2 with model parameters consisting of the radius of the depositional area (m), the fossil pollen count, pollen fall speeds (m/s), relative pollen productivity, and the regional vegetation range and its variance–covariance matrix33. The relative pollen productivity values of 10 plant taxa were selected (Supplementary Table 5), and their pollen percentages accounted for ~90% of the pollen assemblages, covering the principal pollen types in the strata. The pollen productivity of tree taxa was obtained from data on the relative pollen productivity of mountain areas in the upper reaches of the Sanggan River in the Nihewan Basin in Guancen, in the same research area; this was because the pollen productivity of tree taxa is close in the same climate area86. The pollen productivity of herbaceous plants is affected by a wide range of climatic factors, and therefore, the relative pollen productivity of herbaceous plants was selected based on integrated research results for northern China87.
Data availability
All data (Supplementary Data 1–4) used in this study are freely available online (https://doi.org/10.6084/m9.figshare.30415381).
References
Cerling, T. et al. Woody cover and hominin environments in the past 6 million years. Nature 476, 51–56 (2011).
Zeller, E. et al. Human adaptation to diverse biomes over the past 3 million years. Science 380, 604–608 (2023).
Behrensmeyer, A. K. Climate change and human evolution. Science 311, 476–478 (2006).
deMenocal, P. B. African climate change and faunal evolution during the Plio-Pleistocene. Earth Planet. Sci. Lett. 220, 3–24 (2004).
deMenocal, P. B. Climate and human evolution. Science 331, 540–541 (2011).
Trauth, M. H. et al. Northern Hemisphere Glaciation, African climate and human evolution. Quat. Sci. Rev. 268, 107095 (2021).
Bar-Yosef, O. & Belfer-Cohen, A. From Africa to Eurasia—early dispersals. Quat. Int. 75, 19–28 (2001).
Dennell, R. & Roebroeks, W. An Asian perspective on early human dispersal from Africa. Nature 438, 1099–1104 (2005).
Antón, S. C., Potts, R. & Aiello, L. C. Evolution of early Homo: an integrated biological perspective. Science 345, 6192 (2014).
Potts, R. Evolution and climate variability. Science 273, 922–923 (1996).
[Article](https://doi.org/10.1126%2Fscience.273.5277.922) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DyaK28XltVCqu70%3D) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Evolution%20and%20climate%20variability&journal=Science&doi=10.1126%2Fscience.273.5277.922&volume=273&pages=922-923&publication_year=1996&author=Potts%2CR)
- Potts, R. Hominin evolution in settings of strong environmental variability. Quat. Sci. Rev. 73, 1–13 (2013).
[Article](https://doi.org/10.1016%2Fj.quascirev.2013.04.003) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Hominin%20evolution%20in%20settings%20of%20strong%20environmental%20variability&journal=Quat.%20Sci.%20Rev.&doi=10.1016%2Fj.quascirev.2013.04.003&volume=73&pages=1-13&publication_year=2013&author=Potts%2CR)
- Blumenthal, S. A. et al. Aridity and hominin environments. Proc. Natl. Acad. Sci. USA 28, 7331–7336 (2017).
[Article](https://doi.org/10.1073%2Fpnas.1700597114) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Aridity%20and%20hominin%20environments&journal=Proc.%20Natl.%20Acad.%20Sci.%20USA&doi=10.1073%2Fpnas.1700597114&volume=28&pages=7331-7336&publication_year=2017&author=Blumenthal%2CSA)
- Caley, T. et al. A two-million-year-long hydroclimatic context for hominin evolution in southeastern Africa. Nature 560, 76–79 (2018).
[Article](https://doi.org/10.1038%2Fs41586-018-0309-6) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC1cXhtlWjs7rN) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=A%20two-million-year-long%20hydroclimatic%20context%20for%20hominin%20evolution%20in%20southeastern%20Africa&journal=Nature&doi=10.1038%2Fs41586-018-0309-6&volume=560&pages=76-79&publication_year=2018&author=Caley%2CT)
- Magill, C. R., Ashley, G. M. & Freeman, K. H. Water, plants, and early human habitats in eastern Africa. Proc. Natl. Acad. Sci. USA 110, 1175–1180 (2013).
[Article](https://doi.org/10.1073%2Fpnas.1209405109) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC3sXhvFertro%3D) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Water%2C%20plants%2C%20and%20early%20human%20habitats%20in%20eastern%20Africa&journal=Proc.%20Natl.%20Acad.%20Sci.%20USA&doi=10.1073%2Fpnas.1209405109&volume=110&pages=1175-1180&publication_year=2013&author=Magill%2CCR&author=Ashley%2CGM&author=Freeman%2CKH)
- Winder, I. C. et al. Evolution and dispersal of the genus Homo: a landscape approach. J. Hum. Evol. 87, 48–65 (2015).
[Article](https://doi.org/10.1016%2Fj.jhevol.2015.07.002) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Evolution%20and%20dispersal%20of%20the%20genus%20Homo%3A%20a%20landscape%20approach&journal=J.%20Hum.%20Evol.&doi=10.1016%2Fj.jhevol.2015.07.002&volume=87&pages=48-65&publication_year=2015&author=Winder%2CIC)
- Timmermann, A. et al. Climate effects on archaic human habitats and species successions. Nature 604, 495–501 (2022).
[Article](https://doi.org/10.1038%2Fs41586-022-04600-9) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB38XpvV2itbg%3D) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Climate%20effects%20on%20archaic%20human%20habitats%20and%20species%20successions&journal=Nature&doi=10.1038%2Fs41586-022-04600-9&volume=604&pages=495-501&publication_year=2022&author=Timmermann%2CA)
- Zan, J. et al. Mid-Pleistocene aridity and landscape shifts promoted Palearctic hominin dispersals. Nat. Commun. 15, 10279 (2024).
[Article](https://doi.org/10.1038%2Fs41467-024-54767-0) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB2cXis1GhtLjI) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Mid-Pleistocene%20aridity%20and%20landscape%20shifts%20promoted%20Palearctic%20hominin%20dispersals&journal=Nat.%20Commun.&doi=10.1038%2Fs41467-024-54767-0&volume=15&publication_year=2024&author=Zan%2CJ)
- Zeller, E. & Timmermann, A. The evolving three-dimensional landscape of human adaptation. Sci. Adv. 10, eadq3613 (2024).
[Article](https://doi.org/10.1126%2Fsciadv.adq3613) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=The%20evolving%20three-dimensional%20landscape%20of%20human%20adaptation&journal=Sci.%20Adv.&doi=10.1126%2Fsciadv.adq3613&volume=10&publication_year=2024&author=Zeller%2CE&author=Timmermann%2CA)
- deMenocal, P. B. Plio-Pleistocene African climate. Science 270, 53–59 (1995).
[Article](https://doi.org/10.1126%2Fscience.270.5233.53) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DyaK2MXosFOms7c%3D) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Plio-Pleistocene%20African%20climate&journal=Science&doi=10.1126%2Fscience.270.5233.53&volume=270&pages=53-59&publication_year=1995&author=deMenocal%2CPB)
- Domínguez-Rodrigo, M. Is the “Savanna Hypothesis” a dead concept for explaining the emergence of the earliest hominins? Curr. Anthropol. 55, 9–81 (2014).
[Article](https://doi.org/10.1086%2F674530) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Is%20the%20%E2%80%9CSavanna%20Hypothesis%E2%80%9D%20a%20dead%20concept%20for%20explaining%20the%20emergence%20of%20the%20earliest%20hominins%3F&journal=Curr.%20Anthropol.&doi=10.1086%2F674530&volume=55&pages=9-81&publication_year=2014&author=Dom%C3%ADnguez-Rodrigo%2CM)
- Leroy, S. A. G., Arpe, K. & Mikolajewicz, U. Vegetation context and climatic limits of the early Pleistocene hominin dispersal in Europe. Quat. Sci. Rev. 30, 1448–1463 (2011).
[Article](https://doi.org/10.1016%2Fj.quascirev.2010.01.017) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Vegetation%20context%20and%20climatic%20limits%20of%20the%20early%20Pleistocene%20hominin%20dispersal%20in%20Europe&journal=Quat.%20Sci.%20Rev.&doi=10.1016%2Fj.quascirev.2010.01.017&volume=30&pages=1448-1463&publication_year=2011&author=Leroy%2CSAG&author=Arpe%2CK&author=Mikolajewicz%2CU)
- Deng, C. L. et al. Magnetostratigraphic age of the Xiantai Paleolithic site in the Nihewan Basin and implications for early human colonization of Northeast Asia. Earth Planet. Sci. Lett. 244, 336–348 (2006).
[Article](https://doi.org/10.1016%2Fj.epsl.2006.02.001) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BD28XjtVOnt7Y%3D) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Magnetostratigraphic%20age%20of%20the%20Xiantai%20Paleolithic%20site%20in%20the%20Nihewan%20Basin%20and%20implications%20for%20early%20human%20colonization%20of%20Northeast%20Asia&journal=Earth%20Planet.%20Sci.%20Lett.&doi=10.1016%2Fj.epsl.2006.02.001&volume=244&pages=336-348&publication_year=2006&author=Deng%2CCL)
- Belmaker, M. & O’Brien, H. Mesowear study of ungulates from the early Pleistocene site of ‘Ubeidiya (Israel) and the implications for early Homo dispersal from Africa. Quat. Int. 480, 66–77 (2018).
[Article](https://doi.org/10.1016%2Fj.quaint.2017.03.052) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Mesowear%20study%20of%20ungulates%20from%20the%20early%20Pleistocene%20site%20of%20%E2%80%98Ubeidiya%20%28Israel%29%20and%20the%20implications%20for%20early%20Homo%20dispersal%20from%20Africa&journal=Quat.%20Int.&doi=10.1016%2Fj.quaint.2017.03.052&volume=480&pages=66-77&publication_year=2018&author=Belmaker%2CM&author=O%E2%80%99Brien%2CH)
- Agustí, J. & Lordkipanidze, D. An alternative scenario for the first human dispersal out of Africa. L’Anthropologie 123, 682–687 (2019).
[Article](https://doi.org/10.1016%2Fj.anthro.2019.102727) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=An%20alternative%20scenario%20for%20the%20first%20human%20dispersal%20out%20of%20Africa&journal=L%E2%80%99Anthropologie&doi=10.1016%2Fj.anthro.2019.102727&volume=123&pages=682-687&publication_year=2019&author=Agust%C3%AD%2CJ&author=Lordkipanidze%2CD)
- Saarinen, J. et al. Pliocene to middle Pleistocene climate history in the Guadix-Baza Basin, and the environmental conditions of early Homo dispersal in Europe. Quat. Sci. Rev. 268, 107132 (2021).
[Article](https://doi.org/10.1016%2Fj.quascirev.2021.107132) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Pliocene%20to%20middle%20Pleistocene%20climate%20history%20in%20the%20Guadix-Baza%20Basin%2C%20and%20the%20environmental%20conditions%20of%20early%20Homo%20dispersal%20in%20Europe&journal=Quat.%20Sci.%20Rev.&doi=10.1016%2Fj.quascirev.2021.107132&volume=268&publication_year=2021&author=Saarinen%2CJ)
- Ochando, J. et al. Palynological investigations in the Orce archaeological zone, early Pleistocene of southern Spain. Rev. Palaeobot. Palynol. 304, 104725 (2022).
[Article](https://doi.org/10.1016%2Fj.revpalbo.2022.104725) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Palynological%20investigations%20in%20the%20Orce%20archaeological%20zone%2C%20early%20Pleistocene%20of%20southern%20Spain&journal=Rev.%20Palaeobot.%20Palynol.&doi=10.1016%2Fj.revpalbo.2022.104725&volume=304&publication_year=2022&author=Ochando%2CJ)
- Messager, E. et al. Palaeoenvironmental reconstruction of Dmanisi site (Georgia) based on palaeobotanical data. Quat. Int. 223, 20–27 (2010).
[Article](https://doi.org/10.1016%2Fj.quaint.2009.12.016) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Palaeoenvironmental%20reconstruction%20of%20Dmanisi%20site%20%28Georgia%29%20based%20on%20palaeobotanical%20data&journal=Quat.%20Int.&doi=10.1016%2Fj.quaint.2009.12.016&volume=223&pages=20-27&publication_year=2010&author=Messager%2CE)
- Shchelinsky, V. E. et al. Early Palaeolithic sites on the Taman Peninsula (Southern Azov Sea region, Russia): Bogatyri/Sinyaya Balka and Rodniki. Quat. Int. 223–224, 28–35 (2010).
[Article](https://doi.org/10.1016%2Fj.quaint.2009.08.017) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Early%20Palaeolithic%20sites%20on%20the%20Taman%20Peninsula%20%28Southern%20Azov%20Sea%20region%2C%20Russia%29%3A%20Bogatyri%2FSinyaya%20Balka%20and%20Rodniki&journal=Quat.%20Int.&doi=10.1016%2Fj.quaint.2009.08.017&volume=223%E2%80%93224&pages=28-35&publication_year=2010&author=Shchelinsky%2CVE)
- Messager, E. et al. Palaeoenvironments of early hominins in temperate and Mediterranean Eurasia: new palaeobotanical data from Palaeolithic key-sites and synchronous natural sequences. Quat. Sci. Rev. 30, 1439–1447 (2011).
[Article](https://doi.org/10.1016%2Fj.quascirev.2010.09.008) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Palaeoenvironments%20of%20early%20hominins%20in%20temperate%20and%20Mediterranean%20Eurasia%3A%20new%20palaeobotanical%20data%20from%20Palaeolithic%20key-sites%20and%20synchronous%20natural%20sequences&journal=Quat.%20Sci.%20Rev.&doi=10.1016%2Fj.quascirev.2010.09.008&volume=30&pages=1439-1447&publication_year=2011&author=Messager%2CE)
- Sugita, S. et al. Landscape openness and pollen records: a simulation approach. Holocene 9, 409–421 (1999).
[Article](https://doi.org/10.1191%2F095968399666429937) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Landscape%20openness%20and%20pollen%20records%3A%20a%20simulation%20approach&journal=Holocene&doi=10.1191%2F095968399666429937&volume=9&pages=409-421&publication_year=1999&author=Sugita%2CS)
- Pearce, E. A. et al. Substantial light woodland and open vegetation characterized the temperate forest biome before Homo sapiens. Sci. Adv. 9, eadi9135 (2023).
[Article](https://doi.org/10.1126%2Fsciadv.adi9135) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Substantial%20light%20woodland%20and%20open%20vegetation%20characterized%20the%20temperate%20forest%20biome%20before%20Homo%20sapiens&journal=Sci.%20Adv.&doi=10.1126%2Fsciadv.adi9135&volume=9&publication_year=2023&author=Pearce%2CEA)
- Broström, A., Gaillard, M. J., Ihse, M. & Odgaard, B. Pollen-landscape relationships in modern analogues of ancient cultural landscapes in southern Swedend a first step towards quantification of vegetation openness in the past. Veg. Hist. Archaeobot. 7, 189–201 (1998).
[Article](https://link.springer.com/doi/10.1007/BF01146193) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Pollen-landscape%20relationships%20in%20modern%20analogues%20of%20ancient%20cultural%20landscapes%20in%20southern%20Swedend%20a%20first%20step%20towards%20quantification%20of%20vegetation%20openness%20in%20the%20past&journal=Veg.%20Hist.%20Archaeobot.&doi=10.1007%2FBF01146193&volume=7&pages=189-201&publication_year=1998&author=Brostr%C3%B6m%2CA&author=Gaillard%2CMJ&author=Ihse%2CM&author=Odgaard%2CB)
- Sugita, S. Theory of quantitative reconstruction of vegetation I: Pollen from large sites REVEALS regional vegetation composition. Holocene 17, 229–241 (2007).
[Article](https://doi.org/10.1177%2F0959683607075837) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Theory%20of%20quantitative%20reconstruction%20of%20vegetation%20I%3A%20Pollen%20from%20large%20sites%20REVEALS%20regional%20vegetation%20composition&journal=Holocene&doi=10.1177%2F0959683607075837&volume=17&pages=229-241&publication_year=2007&author=Sugita%2CS)
- Pop, E. & Bakels, C. Semi-open environmental conditions during phases of hominin occupation at the Eemian Interglacial basin site Neumark-Nord 2 and its wider environment. Quat. Sci. Rev. 117, 72–81 (2015).
[Article](https://doi.org/10.1016%2Fj.quascirev.2015.03.020) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Semi-open%20environmental%20conditions%20during%20phases%20of%20hominin%20occupation%20at%20the%20Eemian%20Interglacial%20basin%20site%20Neumark-Nord%202%20and%20its%20wider%20environment&journal=Quat.%20Sci.%20Rev.&doi=10.1016%2Fj.quascirev.2015.03.020&volume=117&pages=72-81&publication_year=2015&author=Pop%2CE&author=Bakels%2CC)
- Tarasov, P. E. et al. A pollen-based biome reconstruction over the last 3.562 million years in the Far East Russian Arctic-new insights into climate-vegetation relationships at the regional scale. Clim. Past 9, 2759–2775 (2013).
[Article](https://doi.org/10.5194%2Fcp-9-2759-2013) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=A%20pollen-based%20biome%20reconstruction%20over%20the%20last%203.562%20million%20years%20in%20the%20Far%20East%20Russian%20Arctic-new%20insights%20into%20climate-vegetation%20relationships%20at%20the%20regional%20scale&journal=Clim.%20Past&doi=10.5194%2Fcp-9-2759-2013&volume=9&pages=2759-2775&publication_year=2013&author=Tarasov%2CPE)
- ter Braak, C. J. F. & Juggins, S. Weighted averaging partial least squares regression (WA–PLS): an improved method for reconstructing environmental variables from species assemblages. Hydrobiologia 269–270, 485–502 (1993).
[Article](https://link.springer.com/doi/10.1007/BF00028046) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Weighted%20averaging%20partial%20least%20squares%20regression%20%28WA%E2%80%93PLS%29%3A%20an%20improved%20method%20for%20reconstructing%20environmental%20variables%20from%20species%20assemblages&journal=Hydrobiologia&doi=10.1007%2FBF00028046&volume=269%E2%80%93270&pages=485-502&publication_year=1993&author=Braak%2CCJF&author=Juggins%2CS)
- Zhu, R. X. et al. New evidence on the earliest human presence at high northern latitudes in northeast Asia. Nature 431, 559–562 (2004).
[Article](https://doi.org/10.1038%2Fnature02829) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BD2cXnvFCntb0%3D) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=New%20evidence%20on%20the%20earliest%20human%20presence%20at%20high%20northern%20latitudes%20in%20northeast%20Asia&journal=Nature&doi=10.1038%2Fnature02829&volume=431&pages=559-562&publication_year=2004&author=Zhu%2CRX)
Xie, F., Li, J. & Liu, L. Q. Palaeolithic industries in Nihewan Basin. (Huashan Publishing House, Shijiazhuang, 2006).
Xie, F. An archaeological study of the Nihewan at the old stone age. J. Hebei norm. univ. 43, 5–11 (2018).
[Google Scholar](http://scholar.google.com/scholar_lookup?&title=An%20archaeological%20study%20of%20the%20Nihewan%20at%20the%20old%20stone%20age&journal=J.%20Hebei%20norm.%20univ.&volume=43&pages=5-11&publication_year=2018&author=Xie%2CF)
- Wang, F. G. & Liu, L. Q. Excavations of Paleolithic sites in Majuangou, Yangyuan county, Hebei Province, in 2014. Archaeology 11, 3–19 (2023).
[CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB3sXoslCnsw%3D%3D) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Excavations%20of%20Paleolithic%20sites%20in%20Majuangou%2C%20Yangyuan%20county%2C%20Hebei%20Province%2C%20in%202014&journal=Archaeology&volume=11&pages=3-19&publication_year=2023&author=Wang%2CFG&author=Liu%2CLQ)
- Pang, Q. Q. et al. Late Cenozoic ostracod biostratigraphy and Paleolithic culture relic strata in Nihewan Basin and its discussion for environmental evolution. J. Hebei GEO Univ. 40, 13–40 (2017).
[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Late%20Cenozoic%20ostracod%20biostratigraphy%20and%20Paleolithic%20culture%20relic%20strata%20in%20Nihewan%20Basin%20and%20its%20discussion%20for%20environmental%20evolution&journal=J.%20Hebei%20GEO%20Univ.&volume=40&pages=13-40&publication_year=2017&author=Pang%2CQQ)
- Sun, A. J. et al. Mineral variation of Majuangou profile in the Nihewan Basin during the early Pleistocene and its paleoenvironmental significance. J. Salt Lake Res. 28, 18–24 (2020).
[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Mineral%20variation%20of%20Majuangou%20profile%20in%20the%20Nihewan%20Basin%20during%20the%20early%20Pleistocene%20and%20its%20paleoenvironmental%20significance&journal=J.%20Salt%20Lake%20Res.&volume=28&pages=18-24&publication_year=2020&author=Sun%2CAJ)
- Tu, H. et al. 26Al/10Be burial dating of Majuangou sites in Nihewan Basin, northern China. Quat. Sci. Rev. 346, 109086 (2024).
[Article](https://doi.org/10.1016%2Fj.quascirev.2024.109086) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=26Al%2F10Be%20burial%20dating%20of%20Majuangou%20sites%20in%20Nihewan%20Basin%2C%20northern%20China&journal=Quat.%20Sci.%20Rev.&doi=10.1016%2Fj.quascirev.2024.109086&volume=346&publication_year=2024&author=Tu%2CH)
- Xie, F. & Zhu, R. X. Research status of Majuangou site in Nihewan Basin. Stories Relics 6, 3–5 (2008).
[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Research%20status%20of%20Majuangou%20site%20in%20Nihewan%20Basin&journal=Stories%20Relics&volume=6&pages=3-5&publication_year=2008&author=Xie%2CF&author=Zhu%2CRX)
Chen, M. N. Study on the Nihewan Formation. 1–145 (Ocean Press 1988).
Mclauchlan, K. K. et al. Detecting open vegetation in a forested landscape: pollen and remote sensing data from New England, USA. Holocene 17, 1233–1243 (2007).
[Article](https://doi.org/10.1177%2F0959683607085130) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Detecting%20open%20vegetation%20in%20a%20forested%20landscape%3A%20pollen%20and%20remote%20sensing%20data%20from%20New%20England%2C%20USA&journal=Holocene&doi=10.1177%2F0959683607085130&volume=17&pages=1233-1243&publication_year=2007&author=Mclauchlan%2CKK)
Dimiceli, C. et al. MOD44B MODIS/Terra vegetation continuous fields yearly L3 Global 250 m SIN Grid V006. NASA EOSDIS Land Processes DAAC (2015).
Lisiecki, L. E. & Raymo, M. E. A Plio-Pleistocene stack of 57 globally distributed benthic delta 18O records. Paleoceanography 20, 522–533 (2005).
[Google Scholar](http://scholar.google.com/scholar_lookup?&title=A%20Plio-Pleistocene%20stack%20of%2057%20globally%20distributed%20benthic%20delta%2018O%20records&journal=Paleoceanography&volume=20&pages=522-533&publication_year=2005&author=Lisiecki%2CLE&author=Raymo%2CME)
- Sun, Y. B. et al. Seven million years of wind and precipitation variability on the Chinese Loess Plateau. Earth Planet. Sci. Lett. 297, 525–535 (2010).
[Article](https://doi.org/10.1016%2Fj.epsl.2010.07.004) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC3cXhtV2jt7fJ) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Seven%20million%20years%20of%20wind%20and%20precipitation%20variability%20on%20the%20Chinese%20Loess%20Plateau&journal=Earth%20Planet.%20Sci.%20Lett.&doi=10.1016%2Fj.epsl.2010.07.004&volume=297&pages=525-535&publication_year=2010&author=Sun%2CYB)
- Foister, T. I. F. et al. Homo heterogenus: Variability in early Pleistocene Homo environments. Evol. Anthropol. 32, 373–385 (2023).
[Article](https://doi.org/10.1002%2Fevan.22005) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Homo%20heterogenus%3A%20Variability%20in%20early%20Pleistocene%20Homo%20environments&journal=Evol.%20Anthropol.&doi=10.1002%2Fevan.22005&volume=32&pages=373-385&publication_year=2023&author=Foister%2CTIF)
- Bobe, R. & Behrensmeyer, A. K. The expansion of grassland ecosystems in Africa in relation to mammalian evolution and the origin of the genus Homo. Palaeogeogr. Palaeoclimatol. Palaeoecol. 207, 399–420 (2004).
[Article](https://doi.org/10.1016%2Fj.palaeo.2003.09.033) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=The%20expansion%20of%20grassland%20ecosystems%20in%20Africa%20in%20relation%20to%20mammalian%20evolution%20and%20the%20origin%20of%20the%20genus%20Homo&journal=Palaeogeogr.%20Palaeoclimatol.%20Palaeoecol.&doi=10.1016%2Fj.palaeo.2003.09.033&volume=207&pages=399-420&publication_year=2004&author=Bobe%2CR&author=Behrensmeyer%2CAK)
- Stewart, K. M. Environmental change and hominin exploitation of C4-based resources in wetland/savanna mosaics. J. Hum. Evol. 77, 1–16 (2014).
[Article](https://doi.org/10.1016%2Fj.jhevol.2014.10.003) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Environmental%20change%20and%20hominin%20exploitation%20of%20C4-based%20resources%20in%20wetland%2Fsavanna%20mosaics&journal=J.%20Hum.%20Evol.&doi=10.1016%2Fj.jhevol.2014.10.003&volume=77&pages=1-16&publication_year=2014&author=Stewart%2CKM)
- Szymanek, M. & Julien, M. A. Early and middle Pleistocene climate-environment conditions in central Europe and the hominin settlement record. Quat. Sci. Rev. 198, 56–75 (2018).
[Article](https://doi.org/10.1016%2Fj.quascirev.2018.08.021) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Early%20and%20middle%20Pleistocene%20climate-environment%20conditions%20in%20central%20Europe%20and%20the%20hominin%20settlement%20record&journal=Quat.%20Sci.%20Rev.&doi=10.1016%2Fj.quascirev.2018.08.021&volume=198&pages=56-75&publication_year=2018&author=Szymanek%2CM&author=Julien%2CMA)
- Quinn, R. L. et al. Pedogenic carbonate stable isotopic evidence for wooded habitat preference of early Pleistocene tool makers in the Turkana Basin. J. Hum. Evol. 65, 65–78 (2013).
[Article](https://doi.org/10.1016%2Fj.jhevol.2013.04.002) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Pedogenic%20carbonate%20stable%20isotopic%20evidence%20for%20wooded%20habitat%20preference%20of%20early%20Pleistocene%20tool%20makers%20in%20the%20Turkana%20Basin&journal=J.%20Hum.%20Evol.&doi=10.1016%2Fj.jhevol.2013.04.002&volume=65&pages=65-78&publication_year=2013&author=Quinn%2CRL)
- Reed, K. E. Early hominid evolution and ecological change through the African Plio-Pleistocene. J. Hum. Evol. 32, 289–322 (1997).
[Article](https://doi.org/10.1006%2Fjhev.1996.0106) [CAS](https://www.nature.com/articles/cas-redirect/1:STN:280:DyaK2s3hsFWqtw%3D%3D) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Early%20hominid%20evolution%20and%20ecological%20change%20through%20the%20African%20Plio-Pleistocene&journal=J.%20Hum.%20Evol.&doi=10.1006%2Fjhev.1996.0106&volume=32&pages=289-322&publication_year=1997&author=Reed%2CKE)
- Ao, H. et al. Pleistocene magnetochronology of the fauna and Paleolithic sites in the Nihewan Basin: significance for environmental and hominin evolution in North China. Quat. Geochronol. 18, 78–92 (2013).
[Article](https://doi.org/10.1016%2Fj.quageo.2013.06.004) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Pleistocene%20magnetochronology%20of%20the%20fauna%20and%20Paleolithic%20sites%20in%20the%20Nihewan%20Basin%3A%20significance%20for%20environmental%20and%20hominin%20evolution%20in%20North%20China&journal=Quat.%20Geochronol.&doi=10.1016%2Fj.quageo.2013.06.004&volume=18&pages=78-92&publication_year=2013&author=Ao%2CH)
- Negash, E. W. Modern African ecosystems as landscape-scale analogues for reconstructing woody cover and early hominin environments. J. Hum. Evol. 197, 103604 (2024).
[Article](https://doi.org/10.1016%2Fj.jhevol.2024.103604) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Modern%20African%20ecosystems%20as%20landscape-scale%20analogues%20for%20reconstructing%20woody%20cover%20and%20early%20hominin%20environments&journal=J.%20Hum.%20Evol.&doi=10.1016%2Fj.jhevol.2024.103604&volume=197&publication_year=2024&author=Negash%2CEW)
- Ma, D. D. et al. Site formation processes at Cenjiawan (Nihewan Basin, North China): a case study on the structure of the early Pleistocene archaeological record in lakeshore environments. J. Quat. Sci. 38, 488–504 (2023).
[Article](https://doi.org/10.1002%2Fjqs.3487) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Site%20formation%20processes%20at%20Cenjiawan%20%28Nihewan%20Basin%2C%20North%20China%29%3A%20a%20case%20study%20on%20the%20structure%20of%20the%20early%20Pleistocene%20archaeological%20record%20in%20lakeshore%20environments&journal=J.%20Quat.%20Sci.&doi=10.1002%2Fjqs.3487&volume=38&pages=488-504&publication_year=2023&author=Ma%2CDD)
- Lüdecke, T. et al. Dietary versatility of early Pleistocene hominins. Proc. Natl. Acad. Sci. USA 115, 13330–13335 (2018).
[Article](https://doi.org/10.1073%2Fpnas.1809439115) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Dietary%20versatility%20of%20early%20Pleistocene%20hominins&journal=Proc.%20Natl.%20Acad.%20Sci.%20USA&doi=10.1073%2Fpnas.1809439115&volume=115&pages=13330-13335&publication_year=2018&author=L%C3%BCdecke%2CT)
- Bailey, G. N., Reynolds, S. C. & King, G. C. P. Landscapes of human evolution: models and methods of tectonic geomorphology and the reconstruction of hominin landscapes. J. Hum. Evol. 60, 257–280 (2011).
[Article](https://doi.org/10.1016%2Fj.jhevol.2010.01.004) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Landscapes%20of%20human%20evolution%3A%20models%20and%20methods%20of%20tectonic%20geomorphology%20and%20the%20reconstruction%20of%20hominin%20landscapes&journal=J.%20Hum.%20Evol.&doi=10.1016%2Fj.jhevol.2010.01.004&volume=60&pages=257-280&publication_year=2011&author=Bailey%2CGN&author=Reynolds%2CSC&author=King%2CGCP)
- Qin, Z. & Sun, X. Glacial–interglacial cycles and early human evolution in China. Land 12, 1683 (2023).
[Article](https://doi.org/10.3390%2Fland12091683) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Glacial%E2%80%93interglacial%20cycles%20and%20early%20human%20evolution%20in%20China&journal=Land&doi=10.3390%2Fland12091683&volume=12&publication_year=2023&author=Qin%2CZ&author=Sun%2CX)
- Qiu, Y. H. et al. Magnetostratigraphic chronology and paleoenvironmental background of early Pleistocene humans and mammal in North China. Quat. Sci. 38, 1268–1292 (2018).
[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Magnetostratigraphic%20chronology%20and%20paleoenvironmental%20background%20of%20early%20Pleistocene%20humans%20and%20mammal%20in%20North%20China&journal=Quat.%20Sci.&volume=38&pages=1268-1292&publication_year=2018&author=Qiu%2CYH)
- Wei, Q. Banshan paleolithic site from the lower Pleistocene in the Nihewan Basin in northern China. Acta Anthropol. Sin. 13, 223–238 (1994).
[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Banshan%20paleolithic%20site%20from%20the%20lower%20Pleistocene%20in%20the%20Nihewan%20Basin%20in%20northern%20China&journal=Acta%20Anthropol.%20Sin.&volume=13&pages=223-238&publication_year=1994&author=Wei%2CQ)
- Yang, J. X. et al. Environment of the early Pleistocene Banshan Paleolithic Site in the Nihewan Basin, North China. Front. Earth Sci. 10, 830798 (2022).
[Article](https://doi.org/10.3389%2Ffeart.2022.830798) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Environment%20of%20the%20early%20Pleistocene%20Banshan%20Paleolithic%20Site%20in%20the%20Nihewan%20Basin%2C%20North%20China&journal=Front.%20Earth%20Sci.&doi=10.3389%2Ffeart.2022.830798&volume=10&publication_year=2022&author=Yang%2CJX)
- Roach, N. et al. Pleistocene footprints show intensive use of lake margin habitats by Homo erectus groups. Sci. Rep. 6, 26374 (2016).
[Article](https://doi.org/10.1038%2Fsrep26374) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC28Xotlegsb4%3D) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Pleistocene%20footprints%20show%20intensive%20use%20of%20lake%20margin%20habitats%20by%20Homo%20erectus%20groups&journal=Sci.%20Rep.&doi=10.1038%2Fsrep26374&volume=6&publication_year=2016&author=Roach%2CN)
- Kubat, J. et al. Dietary strategies of Pleistocene Pongo sp. and Homo erectus on Java (Indonesia). Nat. Ecol. Evol. 7, 279–289 (2023).
[Article](https://doi.org/10.1038%2Fs41559-022-01947-0) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Dietary%20strategies%20of%20Pleistocene%20Pongo%20sp.%20and%20Homo%20erectus%20on%20Java%20%28Indonesia%29&journal=Nat.%20Ecol.%20Evol.&doi=10.1038%2Fs41559-022-01947-0&volume=7&pages=279-289&publication_year=2023&author=Kubat%2CJ)
- Rausch, L. et al. An integrated reconstruction of the early Pleistocene palaeoenvironment of Homo erectus in the Denizli Basin (SW Turkey). Geobios 57, 77–95 (2019).
[Article](https://doi.org/10.1016%2Fj.geobios.2019.10.003) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=An%20integrated%20reconstruction%20of%20the%20early%20Pleistocene%20palaeoenvironment%20of%20Homo%20erectus%20in%20the%20Denizli%20Basin%20%28SW%20Turkey%29&journal=Geobios&doi=10.1016%2Fj.geobios.2019.10.003&volume=57&pages=77-95&publication_year=2019&author=Rausch%2CL)
- Potts, R. et al. Increased ecological resource variability during a critical transition in hominin evolution. Sci. Adv. 6, eabc8975 (2020).
[Article](https://doi.org/10.1126%2Fsciadv.abc8975) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB3cXisV2qt7rI) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Increased%20ecological%20resource%20variability%20during%20a%20critical%20transition%20in%20hominin%20evolution&journal=Sci.%20Adv.&doi=10.1126%2Fsciadv.abc8975&volume=6&publication_year=2020&author=Potts%2CR)
- Margari, V. et al. Extreme glacial cooling likely led to hominin depopulation of Europe in the early Pleistocene. Science 381, 693–699 (2023).
[Article](https://doi.org/10.1126%2Fscience.adf4445) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB3sXhs1OrurbN) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Extreme%20glacial%20cooling%20likely%20led%20to%20hominin%20depopulation%20of%20Europe%20in%20the%20early%20Pleistocene&journal=Science&doi=10.1126%2Fscience.adf4445&volume=381&pages=693-699&publication_year=2023&author=Margari%2CV)
- Timmermann, A. et al. Past climate change effects on human evolution. Nat. Rev. Earth Environ. 5, 701–716 (2024).
[Article](https://doi.org/10.1038%2Fs43017-024-00584-4) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Past%20climate%20change%20effects%20on%20human%20evolution&journal=Nat.%20Rev.%20Earth%20Environ.&doi=10.1038%2Fs43017-024-00584-4&volume=5&pages=701-716&publication_year=2024&author=Timmermann%2CA)
- Fægri, K. et al. Textbook of pollen analysis. J. Biogeogr. 12, 328 (1989).
[Google Scholar](http://scholar.google.com/scholar_lookup?&title=Textbook%20of%20pollen%20analysis&journal=J.%20Biogeogr.&volume=12&publication_year=1989&author=F%C3%A6gri%2CK)
Tang, L. Y. et al. An illustrated handbook of Quaternary pollen and spores in China. 1–556 (Science Press, Beijing, 2018).
Grimm, E. C. Tilia Version 1.7.16 Software. (Illinois State Museum, Springfield, IL, USA, 2011).
Zheng, Z. et al. East Asian pollen database: modern pollen distribution and its quantitative relationship with vegetation and climate. J. Biogeogr. 41, 1819–1832 (2014).
[Article](https://doi.org/10.1111%2Fjbi.12361) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=East%20Asian%20pollen%20database%3A%20modern%20pollen%20distribution%20and%20its%20quantitative%20relationship%20with%20vegetation%20and%20climate&journal=J.%20Biogeogr.&doi=10.1111%2Fjbi.12361&volume=41&pages=1819-1832&publication_year=2014&author=Zheng%2CZ)
- Fan, B. S. et al. A novel approach for quantitatively distinguishing between anthropogenic and natural effects on paleovegetation. PNAS Nexus 3, pgae135 (2024).
[Article](https://doi.org/10.1093%2Fpnasnexus%2Fpgae135) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=A%20novel%20approach%20for%20quantitatively%20distinguishing%20between%20anthropogenic%20and%20natural%20effects%20on%20paleovegetation&journal=PNAS%20Nexus&doi=10.1093%2Fpnasnexus%2Fpgae135&volume=3&publication_year=2024&author=Fan%2CBS)
WCS/CIESIN. Last of the wild data version 2: global human influence index (HII) Wildlife Conservation Society (WCS) and Center for International Earth Science Information Network (CIESIN). https://sedac.ciesin.columbia.edu/data/set/wildareas–v2–human–influence–index–geographic (2005).
Cao, X. Y. et al. Impacts of the spatial extent of pollen-climate calibration-set on the absolute values, range and trends of reconstructed Holocene precipitation. Quat. Sci. Rev. 178, 37–53 (2017).
[Article](https://doi.org/10.1016%2Fj.quascirev.2017.10.030) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Impacts%20of%20the%20spatial%20extent%20of%20pollen-climate%20calibration-set%20on%20the%20absolute%20values%2C%20range%20and%20trends%20of%20reconstructed%20Holocene%20precipitation&journal=Quat.%20Sci.%20Rev.&doi=10.1016%2Fj.quascirev.2017.10.030&volume=178&pages=37-53&publication_year=2017&author=Cao%2CXY)
- Wang, T. et al. Vegetation cover dynamics on the northeastern Qinghai-Tibet Plateau since late Marine Isotope Stage 3. Quat. Sci. Rev. 318, 108282 (2023).
[Article](https://doi.org/10.1016%2Fj.quascirev.2023.108292) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Vegetation%20cover%20dynamics%20on%20the%20northeastern%20Qinghai-Tibet%20Plateau%20since%20late%20Marine%20Isotope%20Stage%203&journal=Quat.%20Sci.%20Rev.&doi=10.1016%2Fj.quascirev.2023.108292&volume=318&publication_year=2023&author=Wang%2CT)
- Li, X. Y. et al. Modern pollen-vegetation coverage relationships and its application for Holocene vegetation reconstructions in the central Tianshan mountains. Palaeogeogr. Palaeoclimatol. Palaeoecol. 635, 111963 (2024).
[Article](https://doi.org/10.1016%2Fj.palaeo.2023.111963) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Modern%20pollen-vegetation%20coverage%20relationships%20and%20its%20application%20for%20Holocene%20vegetation%20reconstructions%20in%20the%20central%20Tianshan%20mountains&journal=Palaeogeogr.%20Palaeoclimatol.%20Palaeoecol.&doi=10.1016%2Fj.palaeo.2023.111963&volume=635&publication_year=2024&author=Li%2CXY)
- Andreev, R. & Novenko, E. Quantitative pollen-based reconstructions of climate characteristics and forest coverage for Northern Central Siberia: evaluation of different techniques. Quat. Int. 739, 109879 (2025).
[Article](https://doi.org/10.1016%2Fj.quaint.2025.109879) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Quantitative%20pollen-based%20reconstructions%20of%20climate%20characteristics%20and%20forest%20coverage%20for%20Northern%20Central%20Siberia%3A%20evaluation%20of%20different%20techniques&journal=Quat.%20Int.&doi=10.1016%2Fj.quaint.2025.109879&volume=739&publication_year=2025&author=Andreev%2CR&author=Novenko%2CE)
Birks, H. J. B. et al. Data handling and numerical techniques. In: Tracking environmental change using lake sediments. 5, 249–327 (Springer: Dordrecht, The Netherlands, 2012).
Cao, X. Y. et al. Pollen-based quantitative land-cover reconstruction for northern Asia covering the last 40 ka cal BP. Clim Past 15, 1503–1536 (2019).
[Article](https://doi.org/10.5194%2Fcp-15-1503-2019) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Pollen-based%20quantitative%20land-cover%20reconstruction%20for%20northern%20Asia%20covering%20the%20last%2040%E2%80%89ka%E2%80%89cal%E2%80%89BP&journal=Clim%20Past&doi=10.5194%2Fcp-15-1503-2019&volume=15&pages=1503-1536&publication_year=2019&author=Cao%2CXY)
- Li, F. R. et al. Towards quantification of Holocene anthropogenic land-cover change in temperate China: a review in the light of pollen-based REVEALS reconstructions of regional plant cover. Earth Sci. Rev. 203, 103119 (2020).
[Article](https://doi.org/10.1016%2Fj.earscirev.2020.103119) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Towards%20quantification%20of%20Holocene%20anthropogenic%20land-cover%20change%20in%20temperate%20China%3A%20a%20review%20in%20the%20light%20of%20pollen-based%20REVEALS%20reconstructions%20of%20regional%20plant%20cover&journal=Earth%20Sci.%20Rev.&doi=10.1016%2Fj.earscirev.2020.103119&volume=203&publication_year=2020&author=Li%2CFR)
- Trondman, A. K. et al. Pollen-based quantitative reconstructions of Holocene regional vegetation cover (plant-functional types and land-cover types) in Europe suitable for climate modelling. Glob. Chang. Biol. 21, 676–697 (2015).
[Article](https://doi.org/10.1111%2Fgcb.12737) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Pollen-based%20quantitative%20reconstructions%20of%20Holocene%20regional%20vegetation%20cover%20%28plant-functional%20types%20and%20land-cover%20types%29%20in%20Europe%20suitable%20for%20climate%20modelling&journal=Glob.%20Chang.%20Biol.&doi=10.1111%2Fgcb.12737&volume=21&pages=676-697&publication_year=2015&author=Trondman%2CAK)
- Xia, Z. K. The study on the change of ancient lake shore in the Datong-Yangyuan Basin. Geogr. Res. 11, 52–59 (1992).
[Google Scholar](http://scholar.google.com/scholar_lookup?&title=The%20study%20on%20the%20change%20of%20ancient%20lake%20shore%20in%20the%20Datong-Yangyuan%20Basin&journal=Geogr.%20Res.&volume=11&pages=52-59&publication_year=1992&author=Xia%2CZK)
- Fan, B. S. et al. Quantitative reconstruction of the vegetation of the Qinhuangdao area, western coast of Bohai Sea, North China, during the Little Ice Age. Quat. Int. 641, 51–61 (2022).
[Article](https://doi.org/10.1016%2Fj.quaint.2022.02.024) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Quantitative%20reconstruction%20of%20the%20vegetation%20of%20the%20Qinhuangdao%20area%2C%20western%20coast%20of%20Bohai%20Sea%2C%20North%20China%2C%20during%20the%20Little%20Ice%20Age&journal=Quat.%20Int.&doi=10.1016%2Fj.quaint.2022.02.024&volume=641&pages=51-61&publication_year=2022&author=Fan%2CBS)
- Sun, Y. H., Zhang, S. R. & Xu, Q. H. Pollen-based land cover changes reveal temporal and spatial differences of human activity in north-central China during the Holocene. Catena 219, 106620 (2022).
[Article](https://doi.org/10.1016%2Fj.catena.2022.106620) [Google Scholar](http://scholar.google.com/scholar_lookup?&title=Pollen-based%20land%20cover%20changes%20reveal%20temporal%20and%20spatial%20differences%20of%20human%20activity%20in%20north-central%20China%20during%20the%20Holocene&journal=Catena&doi=10.1016%2Fj.catena.2022.106620&volume=219&publication_year=2022&author=Sun%2CYH&author=Zhang%2CSR&author=Xu%2CQH)
Acknowledgements
This research was supported by the National Natural Science Foundation of China (NSFC) (Grant no. 42377439), the National Key Research and Development Programme of China (Grant no. 2023YFF0804600), and NSFC (Grant nos. T2192954, 41877433, 42507605). We would like to express our gratitude to Dr. Guoqiang Ding from Lanzhou University and Dr. Wensheng Zhang from Hebei GEO University for their assistance in collecting samples. We are grateful to Associate Professor Shengrui Zhang from Hebei Normal University and Dr. Zijing She from Nanjing University for their valuable suggestions.
Author information
Authors and Affiliations
Hebei Technology Innovation Center for Remote Sensing Identification of Environmental Change, Hebei Key Laboratory of Environmental Change and Ecological Construction, School of Geographical Sciences, Hebei Normal University, Shijiazhuang, Hebei, China
Baoshuo Fan, Yuecong Li, Jiaxing Yang, Qinghai Xu, Yawen Ge & Bing Li
State Key Laboratory of Lithospheric and Environmental Coevolution, Institute of Geology and Geophysics, Chinese Academy of Sciences, Beijing, China
Baoshuo Fan, Qingzhen Hao, Deke Xu, Chenglong Deng & Houyuan Lu
Hebei Provincial Institute of Cultural Relics and Archaeology, Shijiazhuang, Hebei, China
Fagang Wang & Fei Xie
College of Earth Sciences, Hebei GEO University, Shijiazhuang, Hebei, China
Zhen Zhang
Authors
- Baoshuo Fan
- Yuecong Li
- Fagang Wang
- Jiaxing Yang
- Zhen Zhang
- Qinghai Xu
- Qingzhen Hao
- Yawen Ge
- Bing Li
- Deke Xu
- Fei Xie
- Chenglong Deng
- Houyuan Lu
Contributions
B.F., Y.L., and H.L. conceived the study; Q.X. provided the modern pollen data; F.W. and F.X. provided field materials; B.F., Z.Z., Y.G., and B.L. collected the samples; B.F., J.Y., and Z.Z. treated and identified the pollen samples; B.F. and D.X. performed statistical analyses; B.F., Y.L., H.L., C.D., and Q.H. wrote the text. All authors commented on the interpretation of the results and gave final approval for publication.
Corresponding authors
Correspondence to Yuecong Li, Chenglong Deng or Houyuan Lu.
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Earth and Environment thanks Julien Louys and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editors: Yiming Wang and Alireza Bahadori. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Fan, B., Li, Y., Wang, F. et al. Relatively open vegetation landscapes promoted early Pleistocene hominin evolution. Commun Earth Environ 6, 1014 (2025). https://doi.org/10.1038/s43247-025-02983-x
Received: 24 July 2025
Accepted: 31 October 2025
Published: 17 December 2025
Version of record: 17 December 2025